nuclear RNAi (Nrde) antisense ribosomal siRNA (risiRNA) inheritance of RNAi
piRNA biogenesis Germ granule nucleolar reshaping
nucleolus stress body (NoSB) aging and epigenetics CRISPR
(本页面由英文页面AI自动翻译, 尚未做校准, 请以英文网页内容为准。)
我们的实验室一直专注于秀丽隐杆线虫中小型调控RNA的机制和功能,并发现了核RNAi缺陷(Nrde)途径,一类新的反义核糖体siRNA (risiRNA)和核仁RNAi。我们还研究了piRNA转录和处理的蛋白质机器,并发现了上游序列转录复合体(USTC),piRNA生物合成和染色体分离复合体(PICS)以及一个新的H3K27me3阅读器UAD-2,这对于piRNA转录至关重要。我们检查了生殖颗粒的结构,并鉴定了E-和D-颗粒,这对于小RNA的产生很重要。同时,我们研究了RNAi的跨代遗传机制,并发现核和细胞质RNAi机器都参与了RNAi的遗传。此外,我们对核仁的结构调控感兴趣,并发现rRNA中间体可以重塑核仁。此外,我们开发了使用CRISPR-Cas9技术诱导大基因组片段缺失、染色体易位和倒位的方法。
RNAi的机制在真核生物中广泛保守。利用小RNA靶向致癌和病毒mRNA的初步成功激发了人们对于小RNA可能最终被用于治疗人类疾病的热情。在合理使用小RNA进行治疗之前,了解它们的生物合成、特异性、运输和内源性角色至关重要。 bi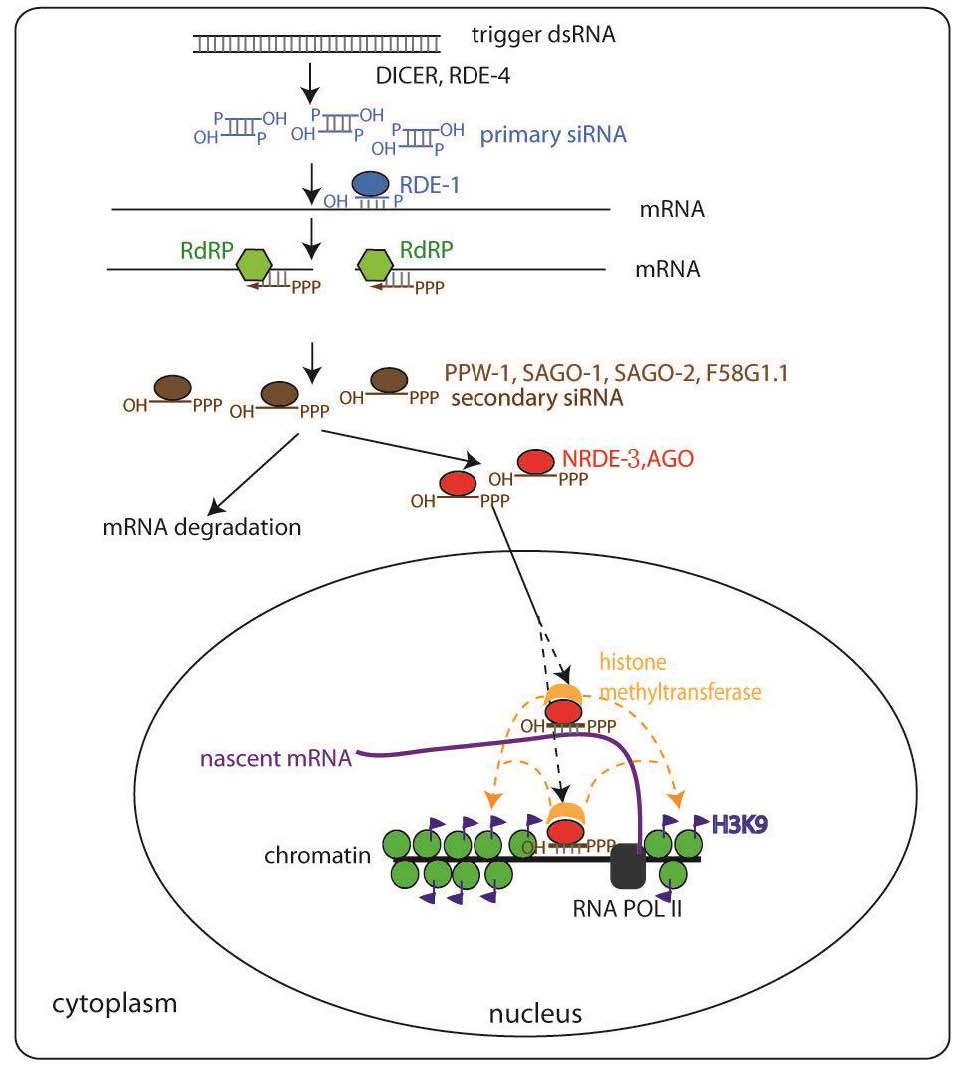 (Science 2008; Nature 2010; PLoS Genetics 2011; Nature Genetics 2012; Genetics 2014; Current Biology 2015)
(Science 2008; Nature 2010; PLoS Genetics 2011; Nature Genetics 2012; Genetics 2014; Current Biology 2015)
(1) 在《C. elegans》中鉴定核RNAi。
我们对小RNA是如何运输和调控的,以及它们在后生动物核中的功能非常感兴趣。为了解决这些问题,我们进行了遗传筛选,以鉴定在线虫模型生物C. elegans中需要核RNAi的因子,并鉴定了三个新基因,称为核RNAi缺陷(NRDE)-1/2/3。我们还鉴定了核和细胞质RNAi途径的分叉,一条新的小RNA运输途径,以及一种新的核基因沉默机制。
NRDE-3是一种Argonaute蛋白。已知Argonaute蛋白可以结合小RNA,通过Watson-Crick碱基配对识别互补的细胞RNA,并通过多种机制抑制基因表达。我们的工作揭示了这些Argonaute蛋白还可以护送小RNA到它们不同的亚细胞室来沉默靶基因。在没有小RNA的情况下,NRDE-3存在于细胞质中。NRDE-3将小RNA从细胞质运输到核,并与RNA聚合酶II产生的新生转录物相关联。这是首次证明特定的Argonaute蛋白除了沉默活性外还具有运输活性。
(2) 核RNAi介导的过早转录终止。
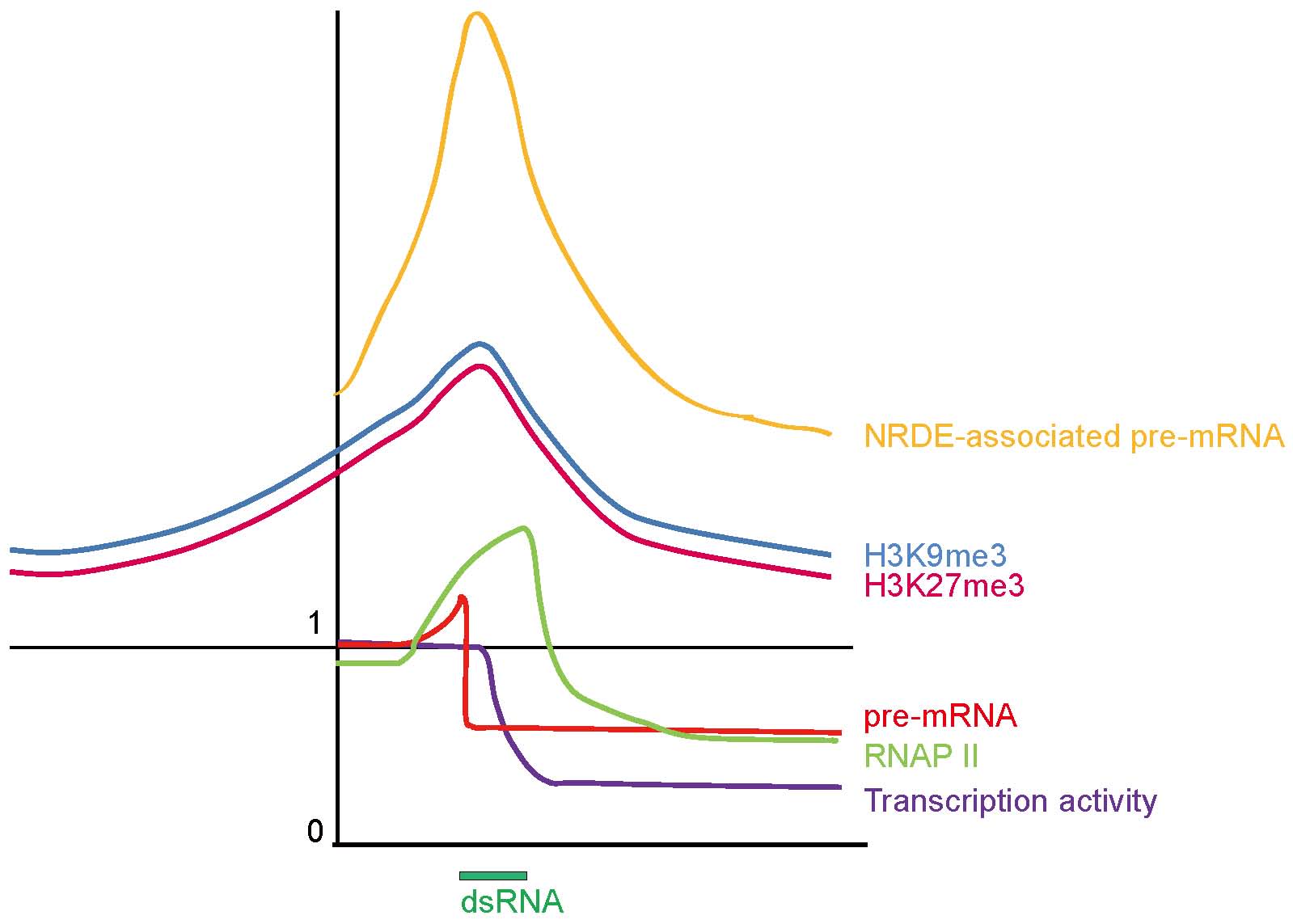 我们的工作还建立了一条新的核基因沉默途径,在这条途径中,小RNA与这些新鉴定的NRDE基因一起作用,通过在RNAi位点的3'端暂停聚合酶来终止RNA聚合酶II介导的转录。通过结合各种遗传和生化测定,我们发现小RNA和NRDE依赖的前mRNAs在RNAi位点的3'端沉默,RNA聚合酶II在RNAi靶向的基因组区域积累,以及RNA聚合酶II占据和转录活性在RNAi位点的3'端减少。这些实验表明,后生动物小RNA引发一个共转录基因沉默程序,并可以作为RNA聚合酶II的反式终止子。
我们的工作还建立了一条新的核基因沉默途径,在这条途径中,小RNA与这些新鉴定的NRDE基因一起作用,通过在RNAi位点的3'端暂停聚合酶来终止RNA聚合酶II介导的转录。通过结合各种遗传和生化测定,我们发现小RNA和NRDE依赖的前mRNAs在RNAi位点的3'端沉默,RNA聚合酶II在RNAi靶向的基因组区域积累,以及RNA聚合酶II占据和转录活性在RNAi位点的3'端减少。这些实验表明,后生动物小RNA引发一个共转录基因沉默程序,并可以作为RNA聚合酶II的反式终止子。
总的来说,这项研究表明后生动物使用一种与A. thaliana和S. pombe不同的机制在核中沉默基因表达。进一步了解小RNA通过NRDEs在核中的功能可能允许更稳定和特异性的基因表达抑制,并促进基础研究和治疗学的进步。
(3) 核RNAi诱导的表观遗传修饰。
由Argonaute样蛋白和小调控RNA组成的核糖核蛋白复合体在广泛的生物学过程中发挥作用。许多小调控RNA被认为至少部分在核内发挥作用,调节转录和修饰染色质。我们表明dsRNA以序列依赖的方式触发H3K27三甲基化,这种修饰可以维持并遗传给后代多代。这种修饰需要秀丽隐杆线虫中的Nrde途径。内源性小RNA,包括但不限于与NRDE-3和HRDE-1相关的内源性siRNA,以NRDE依赖的方式诱导H3K27me3。小RNA介导的H3K9me3和H3K27me3具有不同的遗传要求,在RNAi中发挥不同的作用。H3K27甲基转移酶_mes-2_可能参与小RNA诱导的H3K27me3。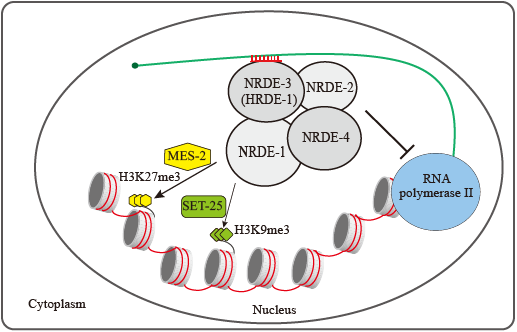
Antisense ribosomal siRNA (risiRNA) and nucleolar RNAi
长期以来,核糖体RNA的小片段被研究界广泛认为是非特异性降解产物,并被视为垃圾序列。关于它们的生物过程、功能和调控机制知之甚少。我们的团队发现一类新的反义核糖体siRNA (risiRNAs)通过核仁RNAi途径下调前rRNA。有趣的是,核仁RNAi途径还涉及NRDE因子,这些因子对核RNAi至关重要。 u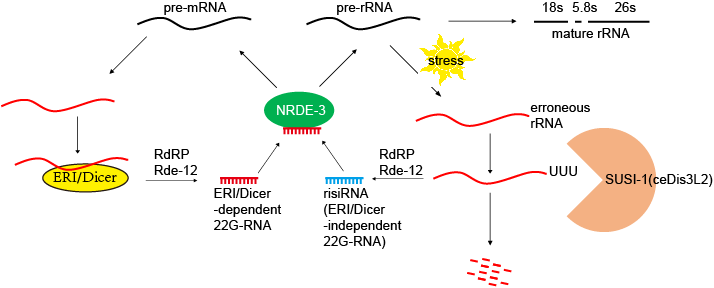 lat (Nature Structural & Molecular Biology 2017; PNAS 2018; Nucleic Acids Research 2021)
lat (Nature Structural & Molecular Biology 2017; PNAS 2018; Nucleic Acids Research 2021)
risiRNAs对环境刺激和基因突变敏感。我们进行了正向和反向遗传筛选,以寻找更多的siRNA抑制因子(susi)突变体,并分离出许多从酵母到人类广泛保守的基因,这些基因参与前rRNA的修饰和处理。这些基因的突变导致rRNA修饰缺陷,并引发risiRNAs的积累,进一步触发细胞质到核和细胞质到核仁的Argonaute蛋白NRDE-3的转位。当涉及rRNA降解的基因SUSI-1(ceDis3L2)和外泌体发生突变时,risiRNAs也显著增加。有趣的是,这种SUSI-1(ceDis3L2)突变也被发现在一种称为Perlman综合征的人类疾病中。因此,我们得出结论,错误的rRNA可以触发risiRNA的产生,随后,打开核RNAi介导的基因沉默途径,以抑制前rRNA表达,这可能提供了一种质量控制机制,以维持rRNA的稳态。
在risiRNAs存在的情况下,NRDE-2和NRDE-3都在核仁中积累,并与RNA聚合酶I共定位,并与前rRNA相关联。risiRNAs通过减少RNAP I占据下游的RNA聚合酶I的转录延伸来抑制RNA聚合酶I的转录延伸。
The mechanism of transgenerational inheritance of RNAi.
RNAi诱导的基因沉默是可遗传的,并能在秀丽隐杆线虫中持续多代,即使在最初的诱导后也是如此。然而,父代获得的RNAi特定性状信息是如何遗传给后代的机制尚不完全清楚。我们鉴定了一种细胞质Argonaute蛋白,WAGO-4,它对RNAi的遗传是必要的。WAGO-4在早期胚胎发育期间不对称地转移到生殖系,在生殖系的核周焦点积累,并且对靶向生殖系和体细胞表达基因的外源RNAi的遗传是必需的。WAGO-4与22G-RNAs及其mRNA靶标结合。有趣的是,与WAGO-4相关的内源性22G-RNAs靶向与CSR-1相同的生殖系基因队列,并在3'端含有未模板化的尿嘧啶添加。聚(U)聚合酶CDE-1需要对22G-RNAs进行未模板化的尿嘧啶化和RNAi的遗传。因此,我们得出结论,除了核RNAi途径外,细胞质RNAi机器也促进RNAi遗传。Rn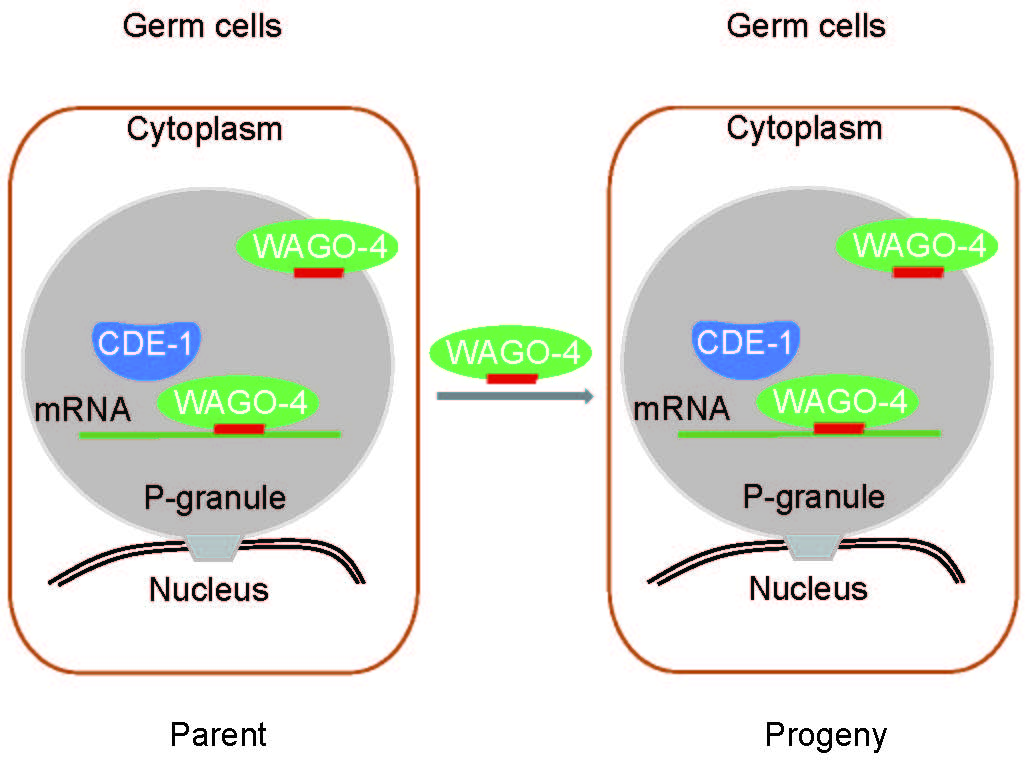 s (Nature Genetics 2012; Cell Reports 2018)
s (Nature Genetics 2012; Cell Reports 2018)
The mechanism of piRNA biogenesis
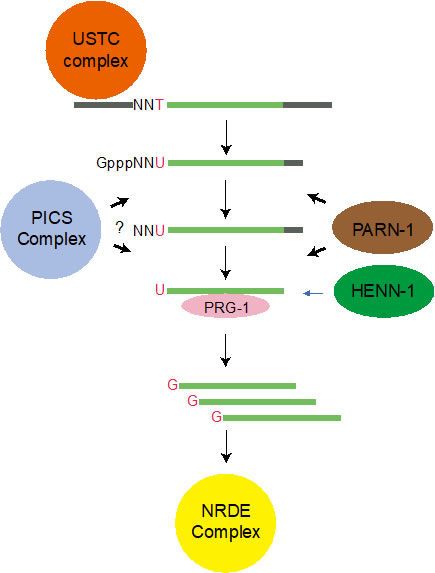
Piwi相互作用RNA (piRNAs)与Piwi蛋白结合,抑制转座子和非自身核酸,维持基因组完整性,对多种生物的生育能力至关重要。在秀丽隐杆线虫中,大多数piRNA前体是从包含数千个单独piRNA转录单元的两个基因组簇中转录而来的。虽然一些基因已被证明需要piRNA生物合成,但piRNA转录的机制仍然难以捉摸。我们使用正向遗传筛选和功能性蛋白质组学方法鉴定了一个上游序列转录复合体(USTC),一个piRNA和染色体分离(PICS)复合体和一个新H3K27me3阅读器UAD-2,这些对piRNA生物合成至关重要。 (Genes & Development 2019; Cell Reports 2019; PNAS 2021; Nature Communications 2021, 2023; Journal of Genetics and Genomics 2022)
(1) piRNA transription
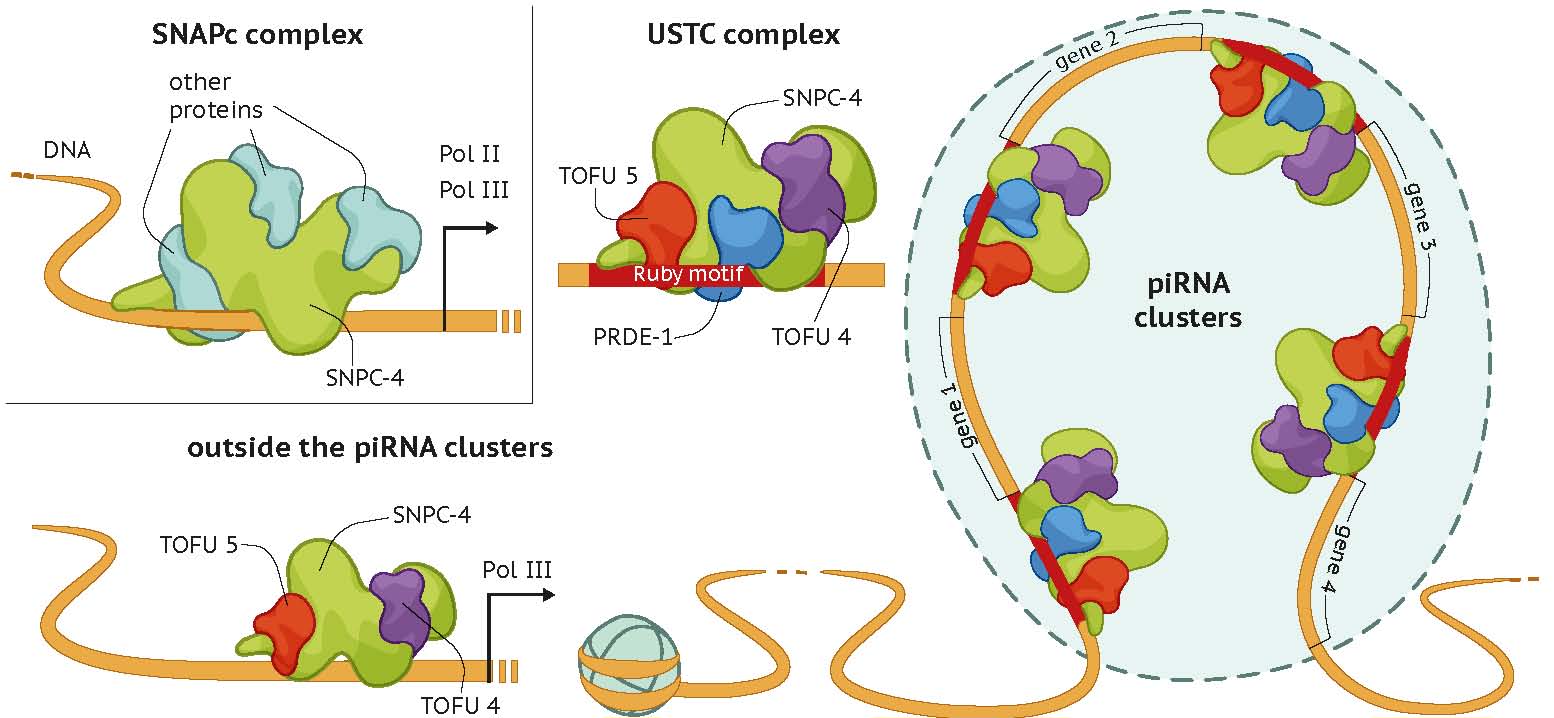
USTC复合体包含PRDE-1、SNPC-4、TOFU-4和TOFU-5。USTC复合体在生殖细胞核中形成一个独特的piRNA焦点,并涂覆piRNA簇基因组位点。USTC因子与I型piRNA基因上游的Ruby基序相关联。
我们进行了正向遗传筛选,并鉴定了UAD-2,这是piRNA焦点形成所必需的。在没有组蛋白3赖氨酸27甲基化或适当的染色质重塑状态下,UAD-2从piRNA焦点中耗尽。UAD-2招募上游序列转录复合体(USTC),该复合体与piRNA启动子结合的Ruby基序,并促进piRNA产生。反之,USTC复合体需要UAD-2与piRNA焦点相关联。因此,异染色质小RNA源位点的转录依赖于组蛋白标记的阅读器和核心转录机器到DNA的协调招募。
(2) piRNA maturation
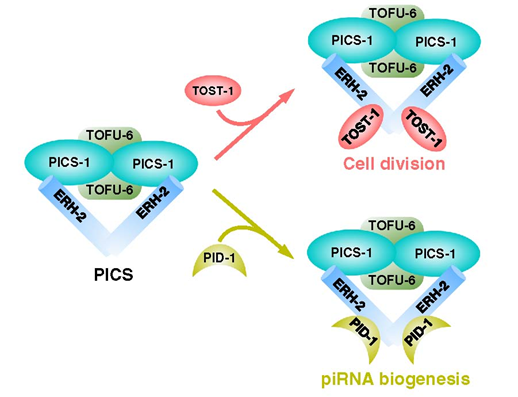
我们使用秀丽隐杆线虫中的功能性蛋白质组学,并鉴定了一个piRNA生物合成和染色体分离(PICS)复合体。PICS复合体包含TOFU-6、PID-1、PICS-1、TOST-1和ERH-2,它们在不同的亚细胞室中表现出动态定位。在生殖细胞中,PICS复合体包含TOFU-6/PICS-1/ERH-2/PID-1,主要集中于核周颗粒区,并参与piRNA处理。在胚胎发育过程中,TOFU-6/PICS-1/ERH-2/TOST-1复合体在核中积累,并在染色体分离中发挥重要作用。这些因子在介导染色体分离中的功能与piRNA产生无关。我们推测PICS因子的不同组成可能帮助细胞协调不同的细胞过程。
我们绘制了PICS亚基之间的相互作用网络,然后通过解决几个复合体结构,包括TOFU-6/PICS-1、ERH-2/PICS-1和ERH-2/TOST-1,揭示了PICS亚基之间相互作用的机制。我们的生化实验还表明PICS作为一个八聚体存在,包含每个亚基的两副本。结合结构分析和突变实验,我们鉴定了PICS亚基界面残基,这些残基对于维持体外完整的PICS复合体至关重要。此外,使用遗传学、细胞生物学和成像实验,我们发现那些破坏PICS内部相互作用网络的突变体,也导致PICS在体内的功能障碍,包括PICS的错位和piRNA水平的降低或异常的染色体分离和细胞分裂。
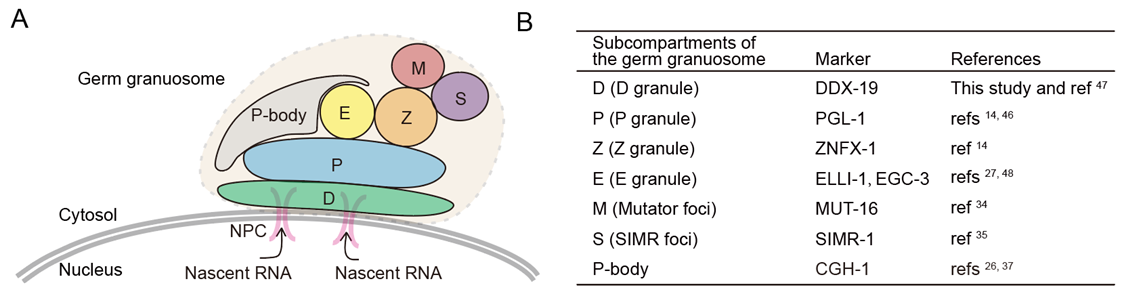
生殖颗粒是存在于大多数动物生殖细胞中的生物分子凝聚体。生殖颗粒的一个功能是通过组织mRNA调控机器,包括基于小RNA的基因调控途径,帮助维持生殖细胞的全能性。秀丽隐杆线虫的生殖颗粒被分隔成多个亚室,其生物学功能大多未知。在这里,我们鉴定了秀丽隐杆线虫生殖颗粒的一个未知亚室,我们称之为E颗粒。E颗粒在生殖颗粒内是非随机定位的。我们鉴定了五种蛋白质,它们定位于E颗粒,包括RNA依赖的RNA聚合酶(RdRP) EGO-1,Dicer相关的解旋酶DRH-3,含有Tudor结构域的蛋白EKL-1,以及两种内在无序蛋白,EGC-1和ELLI-1。EGO-1定位于E颗粒使其能够合成一类特殊的22G RNAs,这些RNAs专门来源于生殖系表达mRNA的5'区域。E颗粒组装的缺陷引发内源性siRNAs的失调产生,这扰乱了生育能力和RNAi反应。我们的结果定义了秀丽隐杆线虫生殖颗粒的一个独特亚室,并表明生殖颗粒分隔的一个功能是促进特殊类别的小调控RNAs的局部产生。(Nature Communications 2024)
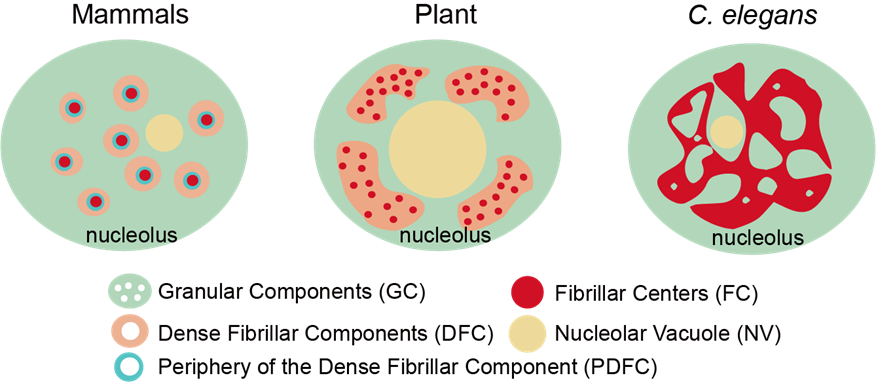
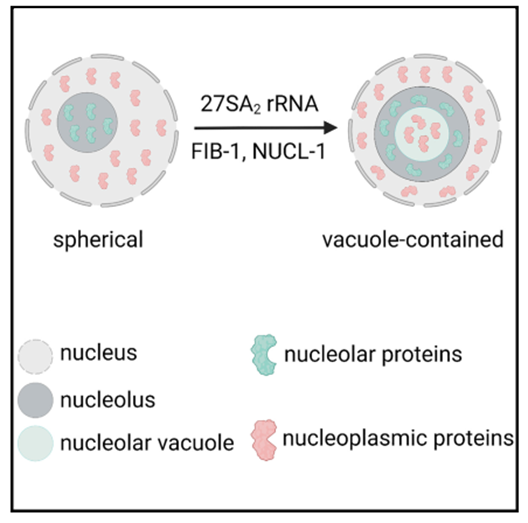
核仁是核内最突出的无膜细胞器。核仁结构是如何调节的尚不清楚。在这里,我们鉴定了秀丽隐杆线虫中的两种核仁类型。I型核仁是球形的,没有可见的核仁空泡(NoVs),rRNA转录和处理因子均匀分布在整个核仁中。II型核仁包含空泡,rRNA转录和处理因子专门积累在周边边缘。NoV包含核质蛋白,并能够与核质交换内容。核仁的高阶结构在秀丽隐杆线虫中是动态调节的。忠实的rRNA处理对于禁止NoVs很重要。27SA 2 rRNA处理因子的耗竭导致NoV形成。抑制RNA聚合酶I (RNAPI)转录和耗竭两个保守的核仁因子,核仁蛋白和纤维蛋白,禁止NoV的形成。这一发现提供了一种协调结构维护和基因表达的机制。(Cell Reports 2023)
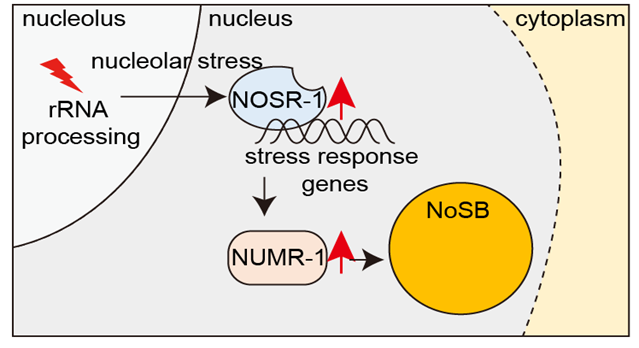
环境刺激不仅改变基因表达谱,还引起细胞结构变化。不同的核体如何响应细胞应激尚不清楚。在这里,我们鉴定了一个名为核仁应激体(NoSB)的亚核细胞器,其形成是由秀丽隐杆线虫中rRNA转录的抑制或rRNA处理和成熟的失活诱导的。NoSB不与其他先前描述的亚核细胞器共定位。我们进行了正向遗传筛选,并鉴定了一个bZIP转录因子,命名为核仁应激反应-1(NOSR-1),这是NoSB形成所必需的。抑制rRNA转录或失活rRNA处理和成熟增加了nosr-1表达。通过使用不同核仁应激条件下的野生型动物和nosr-1突变体的转录组分析,我们鉴定了SR样蛋白NUMR-1(核定位金属响应)是NOSR-1的靶标。有趣的是,NUMR-1是NoSB的一个组成部分,其本身也是NoSB形成所必需的。我们得出结论,NOSR-1/NUMR-1轴可能响应核仁应激,并在秀丽隐杆线虫中介导下游应激反应转录程序和亚核形态变化。(Nature Communications 2024)
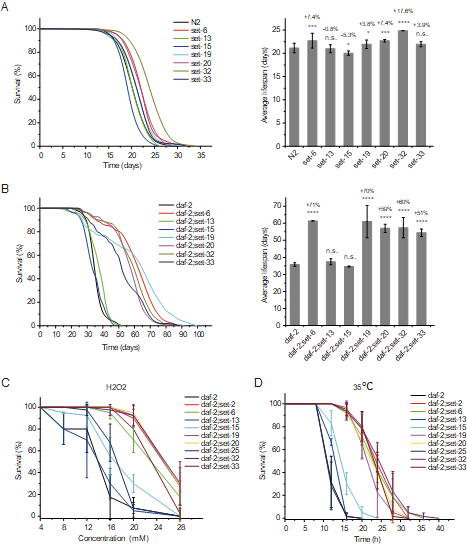
组蛋白甲基化在哺乳动物的发育、基因调控和维持干细胞多能性中发挥着关键作用。最近的研究表明组蛋白甲基化与衰老相关,但潜在的机制尚不清楚。在这项工作中,我们鉴定了一类假定的组蛋白3赖氨酸9单/二甲基转移酶基因(met-2, set-6, set-19, set-20, set-21, set-32和set-33),这些基因的突变在长寿DAF-2(胰岛素生长因子1[IGF-1]受体)突变的秀丽隐杆线虫中诱导协同寿命延长。这些假定的组蛋白甲基转移酶加上daf-2双突变体不仅平均寿命几乎是野生型动物的三倍,最大寿命约为100天,而且还显著增加了对氧化和热应激的抵抗力。协同寿命延长取决于转录因子DAF-16(FOXO)。mRNA-seq实验表明,DAF-16类I基因的mRNA水平,这些基因由DAF-16激活,在daf-2;set双突变体中进一步升高。在这些基因中,tts-1, F35E8.7, ins-35, nhr-62, sod-3, asm-2和Y39G8B.7是daf-2;set-21双突变体寿命延长所必需的。此外,用H3K9me1/2甲基转移酶G9a抑制剂处理daf-2动物也延长了寿命并增加了应激抗性。因此,对DAF-2和H3K9me1/2缺陷介导的协同长寿的研究将有助于更好地理解衰老的分子机制和治疗应用。(eLife 2022; Nature Communications 2023)
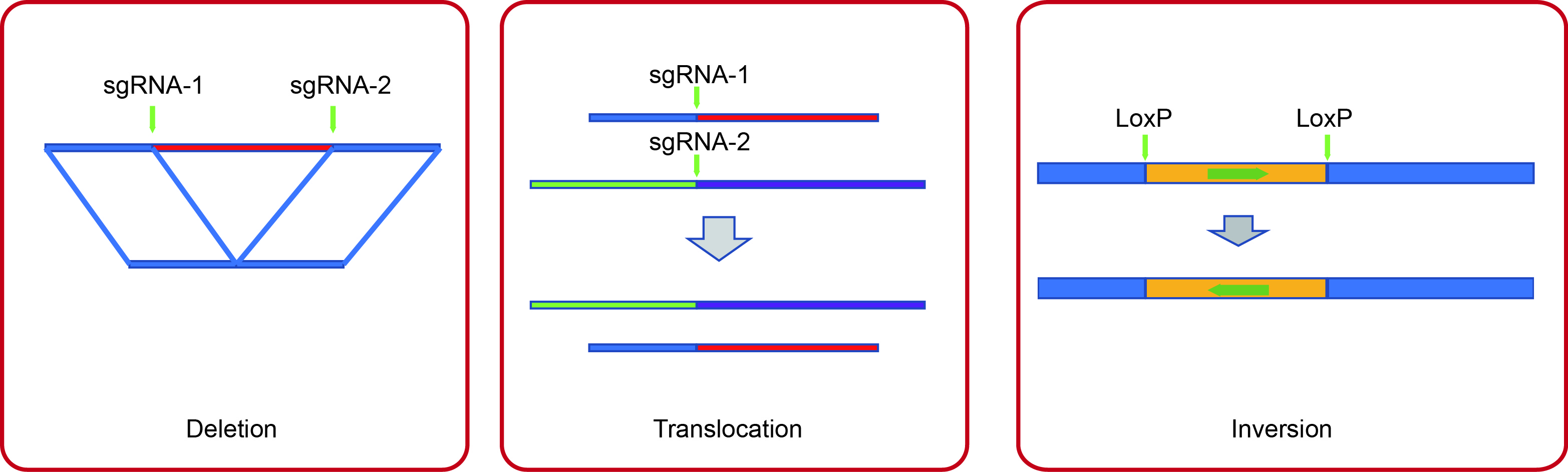
Developing CRISPR/Cas9 technologies for genome manipunation in C. elegans.
CRISPR/Cas9技术已成功应用于基因编辑和染色体工程。在过去的几年中,我们已经开发了CRISPR/Cas9技术用于(a)通过双sgRNAs删除大基因组片段;(b)使用针对不同染色体的sgRNAs诱导染色体易位;(c)通过CRISPR/Cas9和Cre/LoxP手段的组合使用覆盖配对中心的染色体倒位;以及(d)生成一个协议来敲除秀丽隐杆线虫中的基本基因。 (Scientific Reports 2014; Genetics 2015; G3 2018)